The influence of chronic intake of saccharin on rat hepatic and pancreatic function and morphology: gender differences
DOI:
https://doi.org/10.17305/bjbms.2013.2372Keywords:
saccharin, pancrea, liver, glucose, transaminases, ratsAbstract
There are opposite hypotheses on the effect of saccharin. Our aim was reviewing the influence of chronically ingested saccharin on the function and histological structure of liver and pancreas and all this in light of gender differences.
The rats were divided into control group – (Group C) and saccharin-treated group – (Group S) which was given a normal diet and 0.0005% saccharin in drinking water for 6 weeks. Liver and pancreas were histologically processed and quantitative histological analysis was performed. Glucose blood levels and plasma activities of aspartate transaminase (AST) and alanine transaminase (ALT), body weight, and food intake were analyzed.
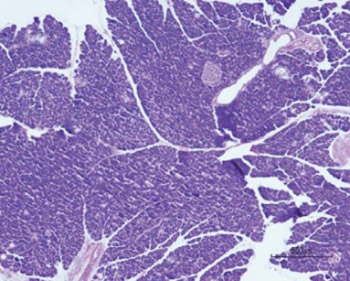
Quantitative histological analysis determined that the values of diameter and volume density of both Langerhans islets and exocrine acini were significantly higher in S group, especially in males. AST levels were significantly higher in treated group. Glucose levels were higher in treated group, mainly due to the values of the female subgroup. Food intake was significantly higher in control group, while weight gain was higher in treated group. Treated males had significantly higher food intake and weight gain in comparison with treated females.
The data presented here suggests that chronic saccharin intake affects the examined parameters. Reported facts reflect various metabolic, hormonal and neural responses in males and females.
Citations
Downloads
Downloads
Additional Files
Published
How to Cite
Accepted 2017-08-17
Published 2013-05-20





