Genetic polymorphisms of ADH1B, ADH1C and ALDH2 in Turkish alcoholics: lack of association with alcoholism and alcoholic cirrhosis
DOI:
https://doi.org/10.17305/bjbms.2015.242Keywords:
alcohol, gene polymorphism, alcoholism, cirrhosis, TurkeyAbstract
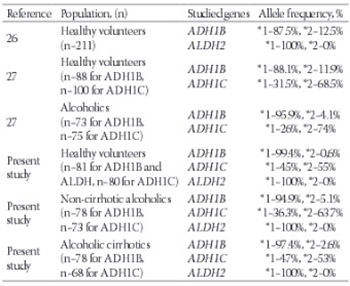
No data exists regarding the alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) gene polymorphisms in Turkish alcoholic cirrhotics. We studied the polymorphisms of ADH1B, ADH1C and ALDH2 genes in alcoholic cirrhotics and compared the results with non-cirrhotic alcoholics and healthy volunteers. Overall, 237 subjects were included for the study: 156 alcoholic patients (78 cirrhotics, 78 non-cirrhotic alcoholics) and 81 healthy volunteers. Three different single-nucleotide-polymorphism genotyping methods were used. ADH1C genotyping was performed using a polymerase chain reaction-restriction fragment length polymorphism method. The identified ADH1C genotypes were named according to the presence or absence of the enzyme restriction sites. ADH1B (Arg47Hys) genotyping was performed using the allele specific primer extension method, and ALDH2 (Glu487Lys) genotyping was performed by a multiplex polymerase chain reaction using two allele-specific primer pairs. For ADH1B, the frequency of allele *1 in the cirrhotics, non-cirrhotic alcoholics and healthy volunteers was 97.4%, 94.9% and 99.4%, respectively. For ADH1C, the frequency of allele *1 in the cirrhotics, non-cirrhotic alcoholics and healthy volunteers was 47%, 36.3% and 45%, respectively. There was no statistical difference between the groups for ADH1B and ADH1C (p>0.05). All alcoholic and non-alcoholic subjects (100%) had the allele *1 for ALDH2. The obtained results for ADH1B, ADH1C, and ALDH gene polymorphisms in the present study are similar to the results of Caucasian studies. ADH1B and ADH1C genetic variations are not related to the development of alcoholism or susceptibility to alcoholic cirrhosis. ALDH2 gene has no genetic variation in the Turkish population.
Citations
Downloads
References
Mizoi Y, Yamamoto K, Ueno Y, Fukunaga T, Harada S. Involvement of genetic polymorphism of alcohol and aldehyde dehydrogenase in individual variation of alcohol metabolism. Alcohol Alcohol 1994;29(6):707–710.
Matsuo K, Wakai K, Hirose K, Ito H, Saito T, Tajima K. Alcohol dehydrogenase 2 His47Arg polymorphism influences drinking habit independently of aldehyde dehydrogenase 2 Glu487Lys polymorphism: analysis of 2,299 Japanese subjects. Cancer Epidemiol Biomarkers Prev 2006;15(5):1009–1013.
http://dx.doi.org/10.1158/1055-9965.EPI-05-0911
Osier M, Pakstis AJ, Kidd JR, Lee JF, Yin SJ, Ko HC, et al. Linkage disequilibrium at the ADH2 and ADH3 loci and risk of alcoholism. Am J Hum Genet 1999;64(4):1147–1157.
http://dx.doi.org/10.1086/302317
Luo HR, Tu GC, Zhang YP. Detection of usual and uncommon aldehyde dehydrogenase alleles by mismatch amplification mutation assay. Clin Chem Lab Med 2001;39(12):1195–1197.
http://dx.doi.org/10.1515/CCLM.2001.189
Macgregor S, Lind PA, Bucholz KK, Hansell NK, Madden PA, Richter MM, et al. Associations of ADH and ALDH2 gene variation with self report alcohol reactions, consumption and dependence: an integrated analysis. Hum Mol Genet 2009;18(3):580–593.
http://dx.doi.org/10.1093/hmg/ddn372
Luo X, Kranzler HR, Zuo L, Wang S, Schork NJ, Gelernter J. Diplotype trend regression analysis of the ADH gene cluster and the ALDH2 gene: multiple significant associations with alcohol dependence. Am J Hum Genet 2006;78(6):973–987.
http://dx.doi.org/10.1086/504113
Roberts-Thomson IC, Butler WJ. Polymorphism and addiction to alcohol. J Gastroenterol Hepatol 2004;19(11):1322–1323.
http://dx.doi.org/10.1111/j.1440-1746.2004.03631.x
Terelius Y, Norsten-Höög C, Cronholm T, Ingelman-Sundberg M. Acetaldehyde as a substrate for ethanol-inducible cytochrome P450 (CYP2E1). Biochem Biophys Res Commun 1991;179(1):689–694.
http://dx.doi.org/10.1016/0006-291X(91)91427-E
Lieber CS. Ethanol metabolism, cirrhosis and alcoholism. Clin Chim Acta 1997;257(1):59–84.
http://dx.doi.org/10.1016/S0009-8981(96)06434-0
Iwahashi K, Ameno S, Ameno K, Okada N, Kinoshita H, Sakae Y, et al. Relationship between alcoholism and CYP2E1 C/D polymorphism. Neuropsychobiology 1998;38(4):218–221.
http://dx.doi.org/10.1159/000026544
Konishi T, Calvillo M, Leng AS, Feng J, Lee T, Lee H, et al. The ADH3*2 and CYP2E1 c2 alleles increase the risk of alcoholism in Mexican American men. Exp Mol Pathol 2003;74(2):183–189.
http://dx.doi.org/10.1016/S0014-4800(03)00006-6
Howard LA, Ahluwalia JS, Lin SK, Sellers EM, Tyndale RF. CYP2E1*1D regulatory polymorphism: association with alcohol and nicotine dependence. Pharmacogenetics 2003;13(6):321–328.
http://dx.doi.org/10.1097/00008571-200306000-00003
Teli MR, Day CP, Burt AD, Bennett MK, James OF. Determinants of progression to cirrhosis or fibrosis in pure alcoholic fatty liver. Lancet 1995;346(8981):987–990.
http://dx.doi.org/10.1016/S0140-6736(95)91685-7
Day CP, Bassendine MF. Genetic predisposition to alcoholic liver disease. Gut 1992;33(11):1444–1447.
http://dx.doi.org/10.1136/gut.33.11.1444
Goedde HW, Agarwal DP, Fritze G, Meier-Tackmann D, Singh S, Beckmann G, et al. Distribution of ADH2 and ALDH2 genotypes in different populations. Hum Genet 1992;88(3):344–346.
http://dx.doi.org/10.1007/BF00197271
Chao YC, Liou SR, Chung YY, Tang HS, Hsu CT, Li TK, et al. Polymorphism of alcohol and aldehyde dehydrogenase genes and alcoholic cirrhosis in Chinese patients. Hepatology 1994;19(2):360–366.
http://dx.doi.org/10.1002/hep.1840190214
Yamauchi M, Maezawa Y, Mizuhara Y, Ohata M, Hirakawa J, Nakajima H, et al. Polymorphisms in alcohol metabolizing enzyme genes and alcoholic cirrhosis in Japanese patients: a multivariate analysis. Hepatology 1995;22(4):1136–1142.
Day CP, Bashir R, James OF, Bassendine MF, Crabb DW, Thomasson HR, et al. Investigation of the role of polymorphisms at the alcohol and aldehyde dehydrogenase loci in genetic predisposition to alcohol-related end-organ damage. Hepatology 1991;14(5):798–801.
http://dx.doi.org/10.1002/hep.1840140509
Chen CC, Lu RB, Chen YC, Wang MF, Chang YC, Li TK, et al. Interaction between the functional polymorphisms of the alcohol-metabolism genes in protection against alcoholism. Am J Hum Genet 1999;65(3):795–807.
http://dx.doi.org/10.1086/302540
Yamauchi M, Maezawa Y, Toda G, Suzuki H, Sakurai S. Association of a restriction fragment length polymorphism in the alcohol dehydrogenase 2 gene with Japanese alcoholic liver cirrhosis. J Hepatol 1995;23(5):519–523.
http://dx.doi.org/10.1016/0168-8278(95)80056-5
Zintzaras E, Stefanidis I, Santos M, Vidal F. Do alcohol-metabolizing enzyme gene polymorphisms increase the risk of alcoholism and alcoholic liver disease? Hepatology 2006;43(2):352–361.
http://dx.doi.org/10.1002/hep.21023
Day CP, James OF, Bassendine MF, Crabb DW, Li TK. Alcohol dehydrogenase polymorphisms and predisposition to alcoholic cirrhosis. Hepatology 1993;18(1):230–232.
http://dx.doi.org/10.1002/hep.1840180140
Chinnaswamy P, Vijayalakshmi V. Subtypes of ADH2 gene in alcoholics. Indian J Clin Biochem 2005;20(2):104–109.
http://dx.doi.org/10.1007/BF02867408
Vidal F, Lorenzo A, Auguet T, Olona M, Broch M, Gutiérrez C, et al. Genetic polymorphisms of ADH2, ADH3, CYP4502E1 Dra-I and Pst-I, and ALDH2 in Spanish men: lack of association with alcoholism and alcoholic liver disease. J Hepatol 2004;41(5):744–750.
http://dx.doi.org/10.1016/j.jhep.2003.06.003
Tamakoshi A, Hamajima N, Kawase H, Wakai K, Katsuda N, Saito T, et al. Duplex polymerase chain reaction with confronting two-pair primers (PCR-CTPP) for genotyping alcohol dehydrogenase beta subunit (ADH2) and aldehyde dehydrogenase 2 (ALDH2). Alcohol Alcohol 2003;38(5):407–410.
http://dx.doi.org/10.1093/alcalc/agg096
Kayaaltı Z, Söylemezoğlu T. Distribution of ADH1B, ALDH2, CYP2E1 *6, and CYP2E1 *7B genotypes in Turkish population. Alcohol 2010;44(5):415–423.
http://dx.doi.org/10.1016/j.alcohol.2010.06.002
Aktas EO, Kocak A, Senol E, Celik HA, Coskunol H, Berdeli A, et al. Determination of the effects of alcohol dehydrogenase (ADH) 1B and ADH1C polymorphisms on alcohol dependence in Turkey. Sci Justice 2012;52(1):58–61.
Downloads
Additional Files
Published
Issue
Section
Categories
How to Cite
Accepted 2014-12-18
Published 2015-05-17





