Hypoxia induces voltage-gated K+ (Kv) channel expression in pulmonary arterial smooth muscle cells through hypoxia-inducible factor-1 (HIF-1)
DOI:
https://doi.org/10.17305/bjbms.2012.2463Keywords:
hypoxia, pulmonary artery smooth muscle cells (PASMCs), voltage-gated K channels, erythropoietin, enhancer hypoxia-inducible factor-1 (HIF-1)Abstract
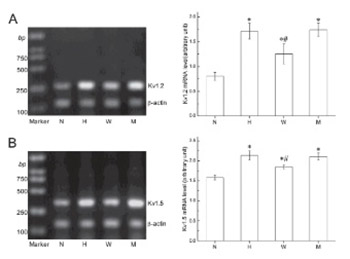
Hypoxia-inducible factor-1 (HIF-1) regulates the expression of hypoxia-inducible genes by binding erythropoietin (EPO) enhancer fragments. Of these genes, HIF-1 upregulates voltage-gated K+1.2 channels (Kv1.2) in rat PC12 cells. Whether HIF-1 regulates hypoxia-induced Kv channel expression in cultured pulmonary artery smooth muscle cells (PASMCs), however, has not been determined. In this study, we investigated the effects of hypoxia on the expression of Kv1.2 Kv1.5, Kv2.1, and Kv9.3 channels in PASMCs and examined the direct role of HIF-1 by transfecting either wild type or mutant EPO enhancer fragments.
Our results showed that 18 h exposure to hypoxia significantly increased the expression of Kv1.2, Kv1.5, Kv2.1, and Kv9.3; and this hypoxia-induced upregulation was completely inhibited after transfection with the wild type but not mutant EPO enhancer fragment.
These results indicate that HIF-1 regulates hypoxia-stimulated induction of Kv1.2, Kv1.5, Kv2.1, and Kv9.3 channels in cultured PASMCs.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
How to Cite
Accepted 2017-09-17
Published 2012-08-20





