Overexpression of miR-361-5p in triple-negative breast cancer (TNBC) inhibits migration and invasion by targeting RQCD1 and inhibiting the EGFR/PI3K/Akt pathway
DOI:
https://doi.org/10.17305/bjbms.2018.3399Keywords:
miR-361-5p, RQCD1, EGFR/PI3K/Akt signaling pathway, cell migration and invasion, triple-negative breast cancer, TNBCAbstract
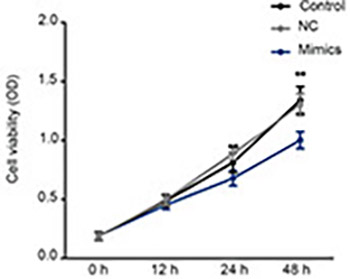
Triple-negative breast cancer (TNBC) is the leading cause of cancer-related death in women. Previous studies indicated that miR-361-5p was downregulated in breast cancer, however, the exact effect of miR-361-5p on TNBC requires further investigation. In the present study, we investigated whether miR-361-5p can act as a tumor suppressor by targeting required for cell differentiation 1 homolog (RQCD1) and inhibiting epidermal growth factor receptor (EGFR)/phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway in TNBC. The expression of miR-361-5p and RQCD1 was determined by quantitative reverse transcription PCR (qRT-PCR) and/or western blot in TNBC and the adjacent tissues. miR-361-5p mimics were constructed and transfected to TNBC cell line MDA-MB-231. Cells were divided into three groups: blank control group, miRNA mimic negative control (NC) group, and miR-361-5p mimics group. Expression of miR-361-5p, mRNA and protein expression of PI3K, Akt, EGFR, phosphorylated (p)-EGFR/PI3K/Akt, and protein expression of RQCD1 and matrix metallopeptidase 9 (MMP-9) in MDA-MB-231 were measured by qRT-PCR/western blot after transfection. Cell viability was determined by CCK-8 assay. Cell migration and invasion ability were evaluated by scratch and transwell assay, respectively. miR-361-5p target gene was determined by bioinformatics analysis and luciferase reporter assay. RQCD1 was identified as a target of miR-361-5p by TargetScan and confirmed by luciferase reporter assay. Downregulated miR-361-5p and upregulated RQCD1 were observed in TNBC tissues. Expression of EGFR, PI3K, Akt and MMP-9 was inhibited in cells treated with miR-361-5p mimics. Transfection of miR-361-5p mimics also inhibited the phosphorylation of EGFR, PI3K, and Akt. Suppressed cell viability, migration, and invasion was found in miR-361-5p mimics groups. Our results indicated that overexpression of miR-361-5p might act as a suppressor in TNBC by targeting RQCD1 to inhibit the EGFR/PI3K/Akt signaling pathway.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
How to Cite
Accepted 2018-05-15
Published 2019-02-12





