Inhibitory effect of microRNA-608 on lung cancer cell proliferation, migration, and invasion by targeting BRD4 through the JAK2/STAT3 pathway
DOI:
https://doi.org/10.17305/bjbms.2019.4216Keywords:
microRNA-608, BRD4, lung cancer, migration, invasionAbstract
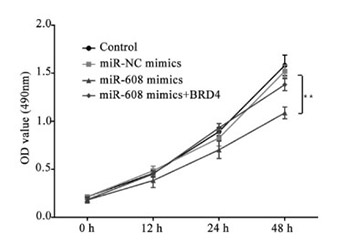
Lung cancer is the leading cause of cancer-related mortality around the world. This malignancy has a 5-year survival rate of 21%, because most of the patients are diagnosed in the middle or late stage of the disease when local metastasis and tumor invasion have already progressed. Therefore, the investigation of the pathogenesis of lung cancer is an issue of crucial importance. MicroRNAs (miRNAs) seem to be involved in the evolution and development of lung cancer. MicroRNA-608 is likely to be downregulated in lung cancer tissues. Regarding this, the current study involved the determination of the fundamental mechanism of microRNA-608 in the development of lung cancer. Based on the results of quantitative reverse transcription polymerase chain reaction (RT-qPCR), the expression level of microRNA-608 was downregulated in 40 lung cancer tissues, compared to that in the adjacent normal tissues. The results of dual-luciferase reporter assay revealed that bromodomain-containing protein 4 (BRD4) was the direct target of microRNA-608. Accordingly, the lung cancer tissues had an elevated expression level of BRD4, in contrast to the adjacent normal tissues. The results of Cell Counting Kit 8 assay demonstrated that the high expression of microRNA-608 notably restrained lung cancer cell proliferation. The scratch wound and transwell assays showed that the upregulation of microRNA-608 suppressed the migration and invasion of lung cancer cells. Finally, the western blot assay showed that in the microRNA-608 mimics group, the expression levels of BRD4, p-JAK2, p-STATA3, CD44, and MMP9 were significantly decreased, compared with those in the negative control miRNA mimics group. Our results indicate that high expression of microRNA-608 inhibits the proliferation, migration, and invasion of lung cancer cells by targeting BRD4 via the JAK2/STAT3 pathway.
Citations
Downloads
Downloads
Additional Files
Published
How to Cite
Accepted 2019-05-09
Published 2020-08-03





