Long noncoding RNA MALAT1 may be a prognostic biomarker in IDH1/2 wild-type primary glioblastomas
DOI:
https://doi.org/10.17305/bjbms.2019.4297Keywords:
Long noncoding RNA, MALAT1, biomarker, prognosis, IDH1/2, primary glioblastoma, isocitrate dehydrogenaseAbstract
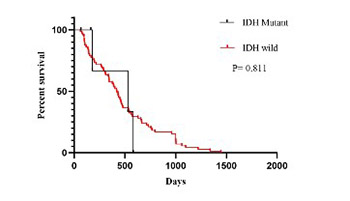
Primary glioblastoma (GB) is the most aggressive type of brain tumors. While mutations in isocitrate dehydrogenase (IDH) genes are frequent in secondary GBs and correlate with a better prognosis, most primary GBs are IDH wild-type. Recent studies have shown that the long noncoding RNA metastasis associated lung adenocarcinoma transcript-1 (MALAT1) is associated with aggressive tumor phenotypes in different cancers. Our aim was to clarify the prognostic significance of MALAT1 in IDH1/2 wild-type primary GB tumors. We analyzed IDH1/2 mutation status in 75 patients with primary GB by DNA sequencing. The expression of MALAT1 was detected in the 75 primary GB tissues and 5 normal brain tissues using reverse transcription quantitative PCR (RT-qPCR). The associations between MALAT1 expression, IDH1/2 mutation status, and clinicopathological variables of patients were determined. IDH1 (R132H) mutation was observed in 5/75 primary GBs. IDH2 (R172H) mutation was not detected in any of our cases. MALAT1 expression was significantly upregulated in primary GB vs. normal brain tissues (p = 0.025). Increased MALAT1 expression in IDH1/2 wild-type primary GBs correlated with patient age and tumor localization (p = 0.032 and p = 0.025, respectively). A multivariate analysis showed that high MALAT1 expression was an unfavorable prognostic factor for overall survival (p = 0.034) in IDH1/2 wild-type primary GBs. High MALAT1 expression may have a prognostic role in primary GBs independent of IDH mutations.
Citations
Downloads
Downloads
Additional Files
Published
How to Cite
Accepted 2019-06-28
Published 2020-02-05





