Association of miRNA-145 with the occurrence and prognosis of hydrosalpinx-induced defective endometrial receptivity
DOI:
https://doi.org/10.17305/bjbms.2020.4538Keywords:
miRNA-145, hydrosalpinx, infertility, inflammation, endometrial cellsAbstract
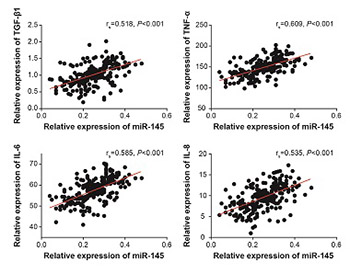
MiR-145 is reported to facilitate inflammation and is also associated with unsuccessful embryonic implantation. Whether miR-145 mediates inflammatory response underlying hydrosalpinx-induced defective endometrial receptivity (ER) remains unclear, and this study attempted to clarify this point. Endometrium samples were collected from hydrosalpinx patients (case, n = 243) and patients with tubal patency/obstruction (control, n = 187). The peripheral blood samples of cases and controls were collected to determine the genotypes of miR-145 SNPs. The value of miR-145 expression in the diagnosis and prognostic estimation of hydrosalpinx was assessed using ROC curve and regression analysis, respectively. Lipopolysaccharide (LPS) cell model was established with endometrial cells, and cells were transfected with miR-145 mimic, inhibitor, or negative control. MiR-145 and cytokine levels were quantified by quantitative reverse transcription PCR or western blot. MiR-145 expression was significantly higher in hydrosalpinx compared to control group, and high miR-145 expression was significantly associated with moderate/severe tube lesion, high pulsatility index (>1.06), and high resistance index (>0.61) in hydrosalpinx patients. ROC curve analysis indicated that monitoring miR-145 expression may be useful for the diagnosis of hydrosalpinx (AUC = 0.704). A alleles of rs41291957 (G>A) and rs353292 (G>A) were significantly associated with an increased risk of hydrosalpinx compared to G allele (p < 0.05), yet the mutant allele of rs353291 (A>G) and rs4705343 (T>C) significantly reduced susceptibility to hydrosalpinx compared to the wild type allele. Treatments with miR-145 mimic and LPS in endometrial cells significantly increased the levels of transforming growth factor-β1, tumor necrosis factor -α, interleukin (IL)-6, and IL-8 compared to negative control, while treatment with miR-145 inhibitor decreased the cytokine levels. In conclusion, abnormally expressed miR-145 may be involved in hydrosalpinx-induced ER defects by regulating the inflammatory response.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
How to Cite
Accepted 2020-03-13
Published 2021-02-01





