ANKRD22 enhances breast cancer cell malignancy by activating the Wnt/β-catenin pathway via modulating NuSAP1 expression
DOI:
https://doi.org/10.17305/bjbms.2020.4701Keywords:
Breast cancer, MDA-MB-415, ANKRD22, NuSAP1, Wnt/β-catenin pathwayAbstract
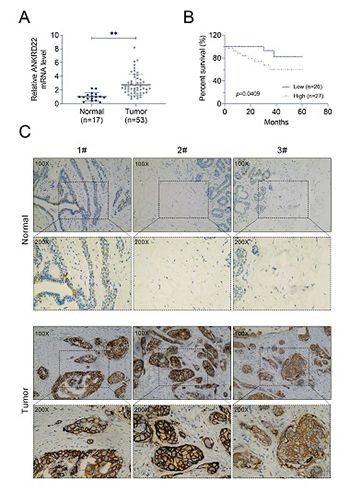
Breast cancer is one of the most prevalent malignancies in women worldwide. Although great advancements have been achieved in the diagnosis and treatment of breast cancer, the prognosis of patients with breast cancer is still poor due to distal recurrence and metastasis after surgery. This study aimed to assess the role of ankyrin repeat domain 22 (ANKRD22) in the progression of breast cancer and investigate the molecular mechanism. Using immunohistochemistry, we demonstrated that the expression level of ANKRD22 in human breast cancer tissues was significantly higher than that in normal breast tissues. ANKRD22 knockdown inhibited the proliferation, invasion, and epithelial-mesenchymal transition (EMT) of breast cancer cells, as confirmed by BrdU, colony formation, transwell, and immunoblot assays. Immunoblot assays further indicated that ANKRD22 regulated the expression of nucleolar and spindle-associated protein 1 (NuSAP1) and then caused the activation of Wnt/β-catenin signaling pathway. Moreover, overexpression of NUSAP1 reversed the inhibitory effects of ANKRD22 knockdown on the proliferation, invasion, and EMT of breast cancer cells. In summary, this study demonstrated that ANKRD22 enhanced breast cancer cell malignancy by activating the Wnt/β-catenin pathway via modulating NuSAP1 expression, which might shed light on new therapeutic approaches for breast cancer.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
How to Cite
Accepted 2020-06-11
Published 2021-06-01





