Astrocytes and human artificial blood-brain barrier models
DOI:
https://doi.org/10.17305/bjbms.2021.6943Keywords:
Astrocytes, blood-brain barrier, in vitro models, bioengineeringAbstract
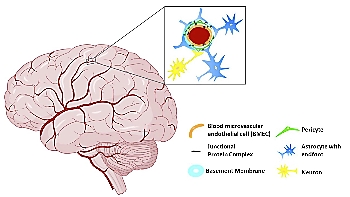
The blood-brain barrier (BBB) functions as a highly selective border of endothelial cells, protecting the central nervous system from potentially harmful substances by selectively controlling the entry of cells and molecules, including components of the immune system. To study the BBB properties, find suitable therapies, and identify new drug targets, there is a need to develop representative in vitro BBB models. In this article, we describe the astrocyte roles in the BBB functioning and human in vitro BBB models.
Citations
Downloads
Download data is not yet available.
Downloads
Additional Files
Published
16-09-2022
License
Copyright (c) 2022 Tanja Zidarič, Lidija Gradišnik, Tomaž Velnar

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
1.
Astrocytes and human artificial blood-brain barrier models. Biomol Biomed [Internet]. 2022 Sep. 16 [cited 2026 Jan. 28];22(5):651-72. Available from: https://www.bjbms.org/ojs/index.php/bjbms/article/view/6943
Received 2022-01-15
Accepted 2022-03-09
Published 2022-09-16
Accepted 2022-03-09
Published 2022-09-16





