Altered fecal microbial and metabolic profile reveals potential mechanisms underlying iron deficiency anemia in pregnant women in China
DOI:
https://doi.org/10.17305/bjbms.2022.7091Keywords:
Iron deficiency anemia, pregnant women, microbiome, metagenomics, metabolic profiling, streptococcus, catecholAbstract
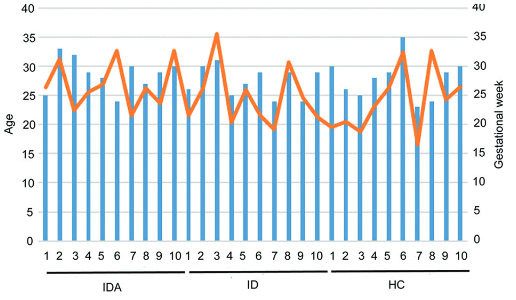
The gut microbiome and its metabolism may provide crucial insight into the cause of iron deficiency anemia (IDA) in pregnant women. This study aimed to investigate the effect of the gut microbiome and its related metabolites on pregnant women with iron deficiency (ID) and IDA. Maternal cubital venous blood and stool samples were collected from healthy control pregnant women (HC, non-anemic, n=10), pregnant women with ID non-anemia (ID, n=10), and IDA (n=10). All groups were subjected to fecal metagenomics and metabolomics. The composition and function of the gut microbiome were then compared in pregnant women with ID and IDA with HC after excluding the possibility of inflammation and insufficient iron absorption capacity. Whole-genome shotgun libraries were prepared by quantifying metagenomic DNA samples with Quant-iT PicoGreen dsDNA Assay. The levels of 41 microbial species, including 21 Streptococci and ten metabolites (catechol), which could serve as siderophores, were increased. In contrast, 3 Bacteroides and six metabolites were decreased in pregnant women with IDA (p<0.05). The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis indicated that the bio-pathways, including biosynthesis of siderophore group non-ribosomal peptides (p<0.01), ABC transporters (p<0.05) and membrane transport of the gut microbiota (p<0.01) in IDA patients were expressed differently compared with HC. Correlation analysis also indicates that these increased bacteria formed strong co-occurring relationships with metabolites in the occurrence and development of IDA in pregnant women. The current study identified that streptococci and catechol (fecal metabolite) were significantly increased in pregnant women with IDA. Therefore, adjusting the intestinal homeostasis using long-term living and eating habits on oral Streptococcus in pregnant women with IDA before iron supplementation may be more conducive to iron supplementation, thus providing novel therapies for IDA.
Citations
Downloads
Downloads
Additional Files
Published
License
Copyright (c) 2022 Haixia Chen, Weigang Wu, Shuming Tang, Rong Fu, Xia Gong, Hu Hou, Junfa Xu

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
Accepted 2022-05-31
Published 2022-10-23





