VHL tumor suppressor as a novel potential candidate biomarker in papillary thyroid carcinoma
DOI:
https://doi.org/10.17305/bjbms.2022.7850Keywords:
Papillary thyroid cancer, PTC, VHL, biomarker, risk stratification, tumor suppressorAbstract
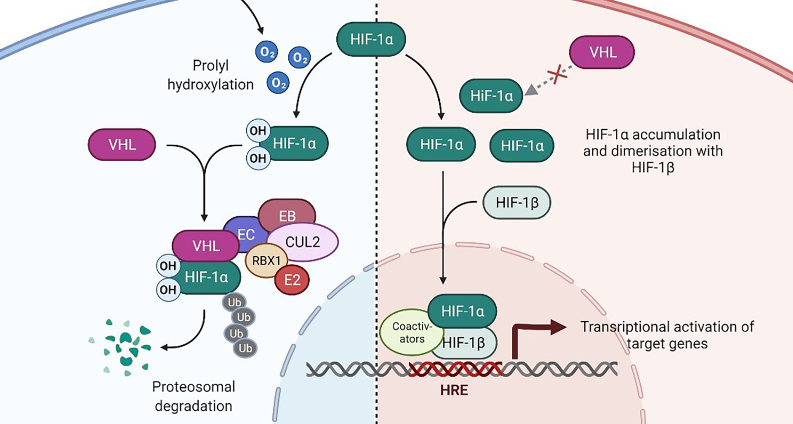
Papillary thyroid carcinoma (PTC) is the most common type of endocrine cancer, with an increasing incidence worldwide. The treatment of PTC is currently the subject of clinical controversy, making it critically important to identify molecular markers that would help improve the risk stratification of PTC patients and optimize the therapeutic approach. The VHL tumor suppressor gene has been implicated in tumorigenesis of various types of carcinoma and linked with their aggressive biological behavior. The role of VHL in the origin and development of PTC has only recently begun to be revealed. In this narrative review we attempt to summarize the existing knowledge that implicates VHL in PTC pathogenesis and to outline its potential significance as a candidate molecular biomarker for the grouping of PTC patients into high and low risk groups.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
License
Copyright (c) 2022 Lidija Todorović, Boban Stanojević

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
Accepted 2022-08-12
Published 2023-01-06





