Effects of salt loading on sympathetic activity and blood pressure in anesthetized two-kidney, one clip hypertensive rats
DOI:
https://doi.org/10.17305/bjbms.2011.2555Keywords:
renovascular hypertension, sodium, angiotensin II, sympathetic nervous systemAbstract
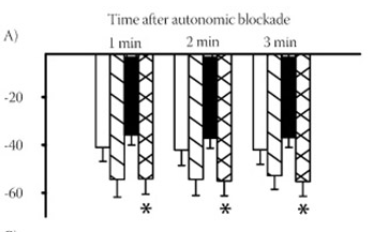
In this study, we investigated the effects of salt loading on sympathetic pressor activity, cardiac autonomic activity, mean arterial pressure (MAP), heart rate (HR) and on the relations between them in anesthetized two-kidney, one clip (2K1C) hypertensive rats. We submitted rats to either renal artery clipping or sham operation. Distilled water or 0.5 % NaCl was given orally to the clipped and sham-operated control rats for 4 weeks. Then, MAP and HR differences between pre- and post- autonomic blockade were evaluated as indexes of sympathetic pressor and cardiac autonomic activity, respectively. The autonomic blockade decreased MAP to the similar levels in all groups (between 81.7±7.6 -87.3±7.1 mmHg). Sympathetic pressor activity was greater in the clipped rats than in its sham-operated controls only under salt loading (55.3±6.2 vs. 37.0±4.1 mmHg, p<0.05). Cardiac autonomic activity was, predominantly, sympathetic and more in the clipped group than in the sham-operated rats under distilled water (48.3±8.6 vs. 19.7±7.0 beats/min, p<0.05) but not under salt loading. Salt loading inverted the relationship between HR and cardiac autonomic activity in 2K1C hypertensive rats (r=-0.76, p=0.046 vs. r=0.89, p=0.019). These results suggest that salt loading may have augmented the effect of renovascular constriction on MAP by affecting the sympathetic pressor activity and the relation between cardiac autonomic activity and HR in 2K1C hypertensive rats.
Citations
Downloads
Additional Files
Published
Issue
Section
Categories
License
Copyright (c) 2017 Bosnian Journal of Basic Medical Sciences

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
Accepted 2017-10-12
Published 2011-11-20





