Exogenous spermidine alleviates diabetic cardiomyopathy via suppressing reactive oxygen species, endoplasmic reticulum stress, and Pannexin-1-mediated ferroptosis
DOI:
https://doi.org/10.17305/bb.2022.8846Keywords:
Diabetic cardiomyopathy, Spermidine, ferroptosis, Pannexin-1, P2X7Abstract
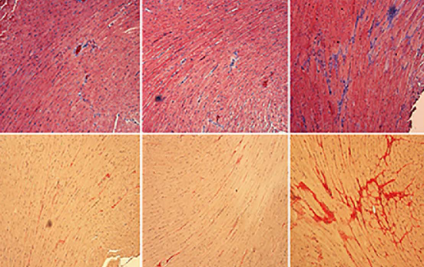
Diabetic cardiomyopathy (DCM) is a serious complication and death cause of diabetes mellitus (DM). Recent cardiology studies suggest that spermidine (SPD) has cardioprotective effects. Here, we verified the hypothesis of SPD’s protective effects on DCM. Therefore, db/db mice and primary neonatal mouse cardiomyocytes were used to observe the effects of SPD. Immunoblotting showed that ornithine decarboxylase (ODC) and SPD/spermine N1-acetyltransferase (SSAT) were downregulated and upregulated in the myocardium of db/db mice, respectively. We found that diabetic mice showed cardiac dysfunction in 12 weeks. Conversely, exogenous SPD could improve cardiac functions and reduce the deposition of collagens, myocardial damage, reactive oxygen species (ROS) levels, and endoplasmic reticulum stress (ERS) in diabetic mouse hearts. Our results also demonstrated that cardiomyocytes displayed ferroptosis and then activated Pannexin-1 expression, which resulted in the increase of the extracellular adenosine triphosphate (ATP). Subsequently, increased ATP as a paracrine molecule combined to purinergic receptor P2X7 to activate ERK1/2 signaling pathway in cardiomyocytes and activated NCOA4-mediated ferroptinophagy to promote lipid peroxidation and ferroptosis. Interestingly, SPD could reverse these molecular processes. Our findings indicate an important new mechanism for DCM and suggest that SPD has potential applicability to protect against deterioration of cardiac function with DCM.
Citations
Downloads
Downloads
Additional Files
Published
Issue
Section
Categories
License
Copyright (c) 2023 Jian Sun, Jiyu Xu, Yong Liu, Yitong Lin, Fengge Wang, Yue Han, Shumin Zhang, Xiaoyan Gao, Changqing Xu, Hui Yuan

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
Accepted 2023-03-11
Published 2023-09-04





